Page 152
are obtained as a result of interspecies hydridization — allopolyploids.
Haploid genome mutations are obtained from the diploid or polyploid forms.
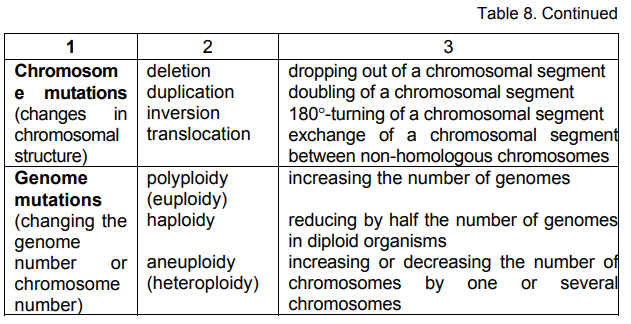
In aneuploid mutations the changes can affect one or several
chromosomes by increasing or reducing their number. Depending on that
they are: trisomia — increasing the genome by one chromosome (2n+1);
tetrasomia — increasing the genome by two chromosomes (2n+2);
monosomia — reducing the genome by one chromosome (2n-1);
nullisomia — reducing the genome by two chromosomes (2n-2), etc.
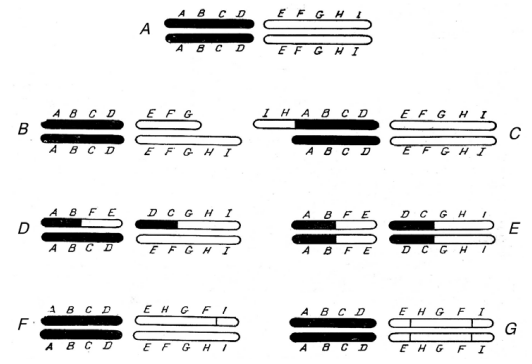
Figure 2–76. Chematic representation of the different types of chromosome mutations (After Rukmansky et al., 1984). A — two normal chromosome pairs; B — deletion; C — duplication; D — heterozygous translocation; E — homozygous translocation; F — heterozygous inversion; G — homozygous inversion
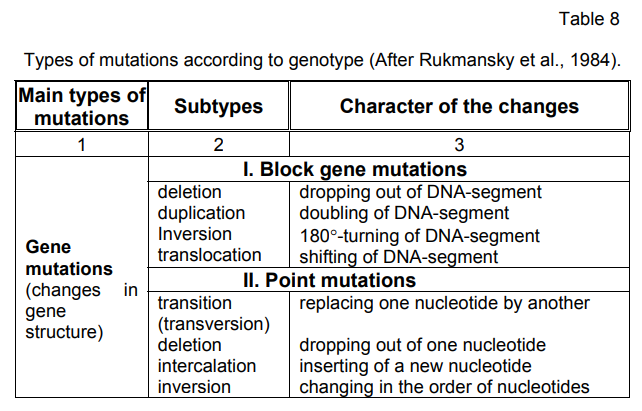
Classification according to phenotype. In accordance with this
classification, mutations are divided into morphological, physiological
and biochemical. Morphological mutations are connected mainly with
changes in the shape and colour. Physiological mutations cause changes
in the physiological processes running in the cells. As to the biochemical
mutations there belong the qualitative and quantitative changes in the
synthesis of certain substances or group of compounds. The latter are well
studied on microorganisms in auxotrophic mutations characterizing by
specific requirements for certain compounds necessary for their growth and
development.