Page 142
image
Figure 2–69. The structural formulas of chlorophyll a and b.
Regardless of the functional differences between mitochondria
and chloroplasts, there is something in common regarding their general
structure: outer and inner membrane of different permeability for ions and
molecules, separated by inter-membrane space; their inner
membranes are located in the matrix and correspondingly in the stroma,
both of them rich in enzymes; they contain specific DNA, RNA and
ribosomes. Schematically that is represented in Figure 2–70.
There are also differences between mitochondria and chloroplasts. Mainly
they lie in the bioprocesses running in them. In chloroplasts the photosynthesis is
realized, which directly depends on the light (i.e. periodically) and as a
result organic compounds are formed and oxygen is released.
Briefly, it is a creative process. In mitochondria the bioprocesses do
not depend on the light, they run continuously, destroying the ready
products of photosynthesis, at that the energy of chemical bonds included in
them is released.
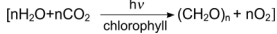
Photosynthesis: CO₂ + H₂O + energy → organic compounds
Respiration: Organic compounds + O₂ → CO₂ + H₂O + energy
As mitochondria (see Fig. 2–66), as well as chloroplasts possess
certain autonomy and can divide independently (Fig. 2–71). These firmly
established facts as well as the numerous data on the presence of specific
DNA, RNA and ribosomes in them were the reasons to accept the old
supposition, that they have endosymbiotic origin. This problem will be
considered in more detail in Section 2.10.