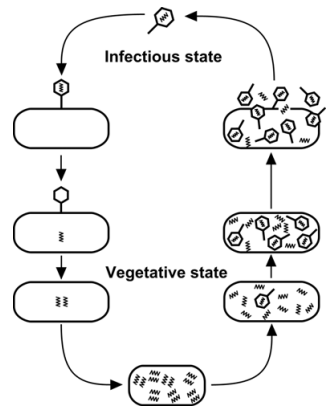
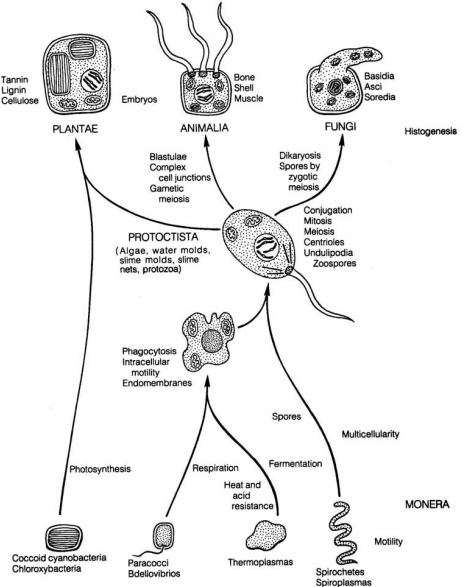
Page 172
image
image
image
image
Figure 2–88. Electron micrographs of some well-known viruses and phages (After Fraenkel-Konrat. 1985) 1 — tobacco mosaic virus (18 x 300 nm); 2 — potato virus X (13 x 600 nm); 3 — turnip yellow mosaic virus (diameter 29 nm); 4 — human herpes virus (about 200 nm diameter). The tubular capsomeres, 162 in all, are clearly distinguishable; 5 — adenovirus (diameter 80 nm) largely composed of hexones, with fibres (27 nm in length) attached to the pentons; 6 — phage φx 174 (diameter 27 nm, with spikes at the vertices of the icosahedron); 7 — phage λ (diameter of the head — 54 nm, tail — 15 x 150 nm); 8 — phage T4 (head — 80—95 nm and tail — 16 x 110 nm) with clearly visible neck, tail fibers, base plate, and spikes.