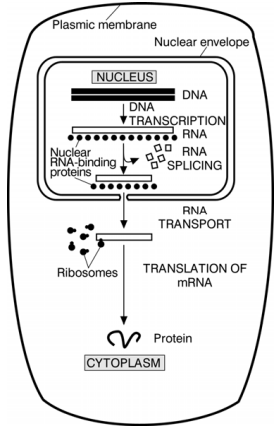
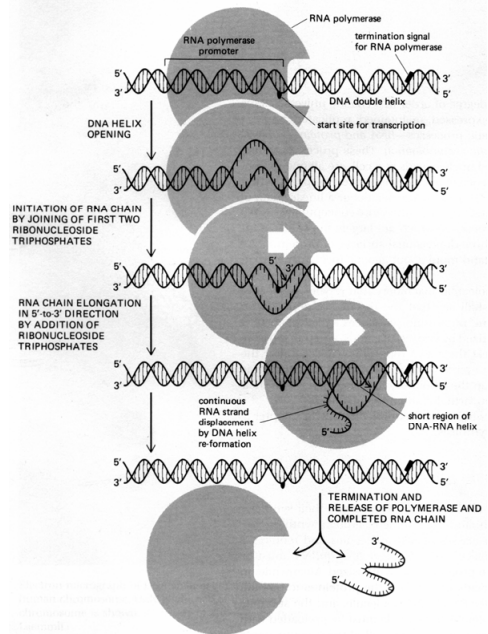
Page 112
image
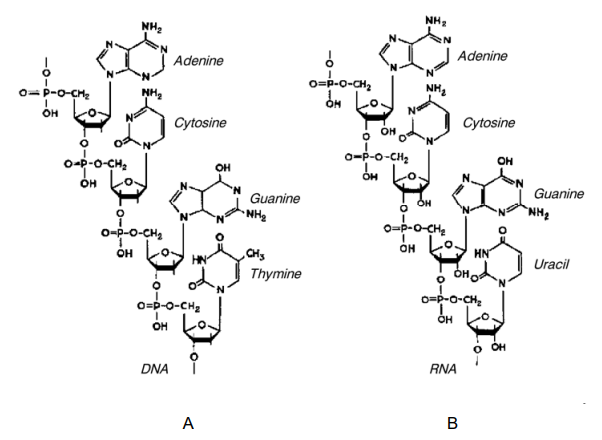
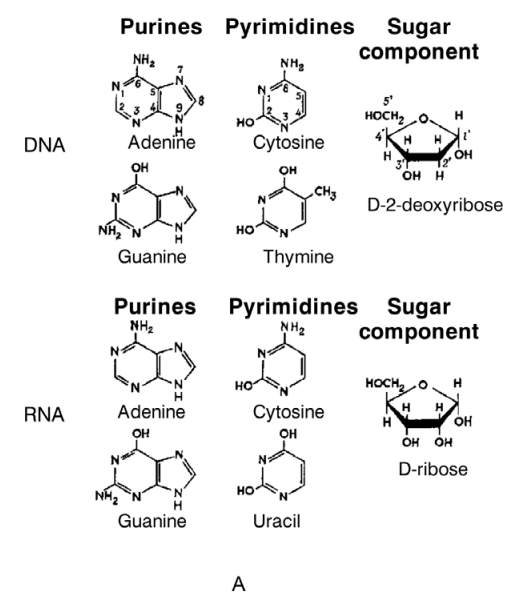
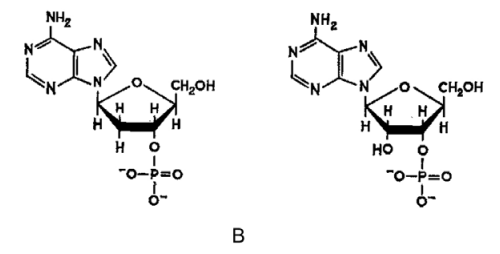
Figure 2–41. Three-dimensional models of the double helices of DNA (A) and RNA (B). Both structures are formed from two antiparallel polynucleotide chains linked by hydrogen bonds between the complementary bases. DNA is presented in B-form, and RNA — in its A form (Courtesy of Sung-How Kim; From Alberts et al. 1986).
The Watson and Crick model places the DNA molecule in a central
position and the protein components in a subordinate one. Since this is
one of the key issues of the modern biology and genetics it cannot be
left out without the due attention.